The herbaceous understory of a forest makes up less than 1% of the biomass yet more than 90% of vascular plant diversity. Photo by Shelby Perry
Biodiversity: The Language of Wilderness
Stewardship
April 2022 | Volume 28, Number 1
What We Know and What We Don’t
I sometimes try to imagine what a northern forest would look like through the eyes of a Black-capped Chickadee. These small birds can see ultraviolet light, which means they can perceive a whole range of colors, textures, and contrasts invisible to us. They can track fast motion much better than we can and they see small objects in more detail, with multiple focal points over a wider field of view. This allows them to see an entire universe of tiny insects, mites, spiders, beetle wings, insect eggs, larvae, and more tucked away in bark crevices, on clusters of dead leaves, and along twigs (Haskell 2012). A chickadee gleans her meals from this smorgasbord of invertebrates jabbing them under flaking bark, stored for later, and is able to remember more than two thousand hiding places at once (Sibley 2020). The richness of a chickadee’s visual world brings home how little we know about the diversity of life in northeastern US forests and the complex relationships that exist among the species living here.
Every northeastern US state has an action plan that documents the status and vulnerability of its native wildlife (defined as any living being that is part of the natural world, including plants, animals, and fungi (Hunter 1990)).These plans focus primarily on species we can readily see and identify—vertebrates, a handful of invertebrate groups such as mussels, butterflies, and beetles, and in some states, vascular plants. We know a fair bit about vertebrates and vascular plants. Maine’s 2015 plan, for example, documents 17 species of reptiles, 18 species of amphibians, 39 species of inland fish, 61 species of non-marine mammals, 423 species of birds, and more than 2,100 species of plants (MDIFW 2015). This is impressive, but also humbling when one stops to consider that mosses, lichens, liverworts, and fungi outnumber vascular plants by more than two to one and vertebrates account for less than two percent of the state’s known wildlife species (MDIFW 2015; Gawler et al. 1996)(Figure 1). Globally, the number of insect species alone—the most diverse group of organisms—exceeds the number of vertebrate species by more than a million (Figure 2). While biologists in Maine are actively cataloging the distribution, status, and life histories of butterflies, dragonflies, bees, and a small handful of other orders of arthropods and mollusks, there are entire kingdoms and phyla about which almost nothing is known, such as protists, sponges, hydras and hydrozoans, flatworms, and nematodes, not to mention bacteria and other microbes. It is these largely inconspicuous species that make up the vast majority of the region’s faunal diversity in terms of both species richness and biomass (Figure 1). In short, if we were to describe the region’s natural history in a book, most of the chapters would be missing (Figure 2).
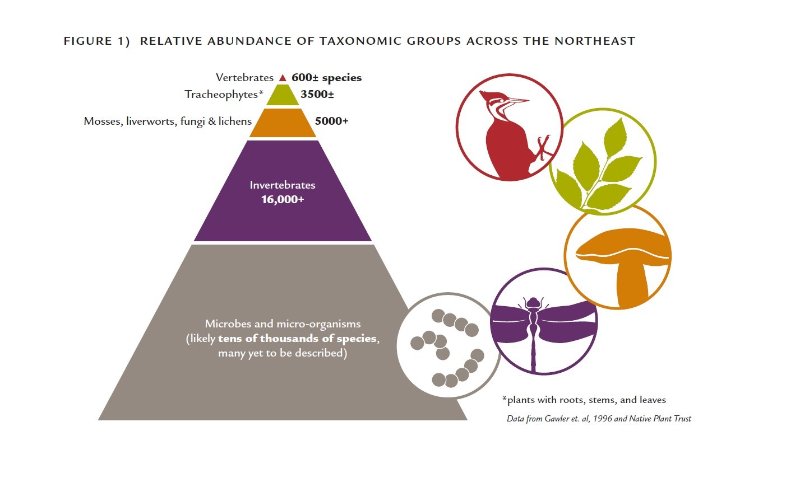
Figure 1 – Relative abundance of taxonomic groups across the Northeast.
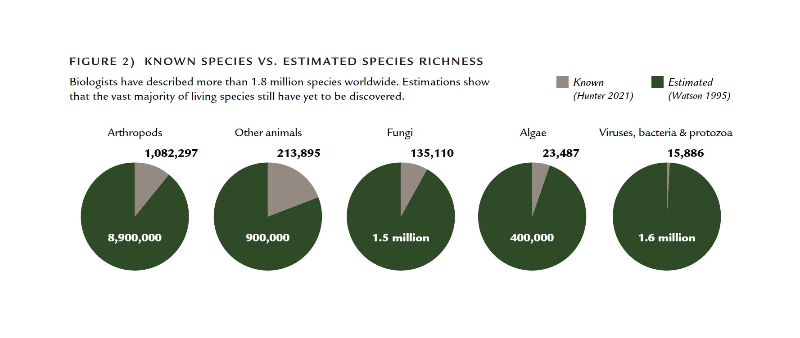
Figure 2 – Known species vs. estimated species richness.
Wilderness and Biodiversity: Disproportionate Benefits
Given how little we know, how do we ensure that the fullest extent of a region’s native biodiversity is protected? It is well documented that intact wilderness areas and landscapes with a light human footprint harbor a greater variety and higher populations of native species than urban and intensively managed landscapes. This is true for lichens, mosses, salamanders, frogs, birds, bats, many herbaceous plants, and even trees (see for example, Lapin 2005; Selva 1996; Whitman and Hagan 2000; Miller et al. 2018; Haney and Schaadt 1996; Zlonis and Niemi 2014). In addition, recent research indicates that wilderness areas support more rare species globally, including those that are found nowhere else (Lapin 2005; Mittermeier et al. 2003). While wilderness areas are critically important for these reasons, they are more than places to count species. Much of the Northeast’s biodiversity is tied to the complex structure of undisturbed forests. Forests that have evolved with little or no human intervention typically have taller and more ragged canopies with more large limbs up high, fewer but larger trees with more cavities and hollows, more and larger dead standing and fallen trees, a higher diversity of lichens, mosses and herbaceous plants, smaller gaps in the canopy, uneven forest floors with dry mounds and damp hollows formed from tip ups, more seeps and intermittent streams, fewer invasive species, deeper litter and humus layers, and a host of other features absent in younger forests (Lapin 2005; Haskell 2017; Maloof 2016). The variety of microclimates on a single old tree is astounding, ranging from moist moss-covered trunk to sunny windswept tree top, to cavities and deeply fissured bark—and this doesn’t include the multitude of microclimates and habitats below ground (Haskell 2017)(Figure 3).

Figure 3 – Forests that are old or minimally impacted by people have a higher occurrence of the following features. Bramhall Wilderness Preserve, pictured above, is a mature forest in Vermont protected by Northeast Wilderness Trust.
We are only beginning to understand how essential the connections and interactions among all of a forest’s inhabitants—from soil to canopy—are to a thriving forest ecosystem. One example is the keystone role of fungi. Mycorrhizal fungi control soil moisture, which in turn impacts wood decomposition rates and nutrient cycling carried out by other fungi, microbes and insects (Simard et al. 2012; Swan and Kominoski 2012). This, in turn, supports extensive food webs, not only in the soil, but in the streams, lakes and wetlands within forested landscapes. Haskell (2017) emphasizes the importance of the “networked nature of trees,” especially dead logs, branches, and roots. These are “focal points for thousands of relationships,” providing food and shelter for more than half the species in a forest. He also notes that soils in northern forests hold more carbon than all the tree trunks, branches, lichens, and other above-ground life combined. We are learning that this intricate tapestry of relationships is a form of communication, with signals in the form of carbon, water, and other chemicals sent back and forth from birch to fir to alder, between mother trees and their offspring, between dying trees and nearby seedlings (Simard 2021).
Another example of the tightly interwoven nature of forest communities is evident in the ground layer. For example, herbaceous plants on the forest floor account for only one percent of plant biomass in temperate forests but encompass ninety-nine percent of vascular plant diversity (Maloof 2016). The diversity of all other organisms in a forest, from butterflies to mammals, is more closely correlated with herbaceous plant diversity than with tree diversity, with herbs providing food for pollinators, herbivores, dispersers, and decomposers (Meier et al. 1995; Maloof 2016). Wildlands ensure that these intricate relationships are not severed and an intact foundation is maintained. They are places where we can learn the language of forests in all its complexity and beauty.

Figure 4 – The visual world of a chickadee illustrates how vast and complex the layers of biodiversity in the northern forest are, beyond what is typically perceived by the human observer. Photo by Larry Master, www.masterimages.org

Figure 6 – Arthropods may receive less conservation attention than charismatic animals like birds, fish, and mammals, but they are just as integral to the web of life. This six-spotted tiger beetle was seen at Northeast Wilderness Trust’s Lone Mountain Wilderness Sanctuary in Maine. Photo by Shelby Perry
Extinction Debt
Today, only about 0.4% of forestland in the US Northeast is old growth (Davis 1996), and in Maine—the most heavily forested state in the region—less than four percent is more than 100 years old, the age at which a stand begins to acquire the complex structure associated with older forests (Whitman and Hagan 2009; 2019 FIA estimate). Before European colonization, two-thirds of the region was old growth, and 55–60% of Maine’s northern forest was more than 150 years old (Lapin 2005; Lorimer 1977).
The disconnect between forests that are managed to reach “financial maturity” (typically 50 to 75 years old) and those allowed to reach biological maturity (~200 years+) has caused a profound ecological shift that we are only beginning to understand. There is growing evidence that economically efficient and intensive management of stands results in significant loss of biodiversity (Hagan and Whitman 2004). This loss may not become apparent until decades after a harvest—a phenomenon known as extinction debt (Hanski 2000). Researchers in Finland estimate that a thousand forest species will be lost there in the next two to three decades due to logging that has already happened (Hagan and Whitman 2004; Hanski 2000) and Tikkanen and others (2006) found that half of the country’s documented endangered species were dependent on old growth forest conditions. The species most at risk are those that move or disperse slowly through the landscape and prefer large standing and fallen trees and deep forest interiors as their habitat. They tend to be the uncharismatic and inconspicuous ones such as mosses, lichens, fungi, vernal forest herbs, and insects. Species loss occurs when there is not enough habitat for long-term persistence and when remaining habitat “islands” are too far apart. Once older forest elements are lost from a stand, it can take centuries for species that depend on those characteristics to return. This is because it takes time for structural features to redevelop, and once they’ve reappeared it takes some time for the species to find them (Hagan and Whitman 2004; Meier et al. 1995). In the face of habitat loss and change, wildlands provide ecological continuity over time. They act as refugia, retaining biological legacies that serve as source areas from which species can recolonize surrounding managed forestland (Hagan and Whitman 2004).

Figure 5 – The diversity of salamander species is greater in wilderness areas and landscapes with a light human footprint. Photo Red Eft by David Middleton
Ecological Amnesia
When I walk through a typical northeastern forest managed for timber, or look out at a sea of trees from a mountaintop, I am usually looking at trees that are 60 or 70 years old at best. To the trained eye, it is hard not to notice what is missing. The top of the forest has essentially been taken off and the canopy that’s left often doesn’t close before the next harvest or grows into a dense thicket of young trees with few gaps from treefalls or other natural disturbances. There are few large standing or fallen dead trees and supercanopy pines are rare. There is no thick moss layer under spruce and hemlock and the seeps that used to course over the forest floor in the spring have been smoothed by repeated logging operations (Figure 3). Plants like marginal fern, Braun’s holly fern, or rattlesnake plantain—once common sights on a rich hardwood forest floor—are now rarely seen and the forest in spring grows quieter year by year. Instead of a collection of distinct places—each different from any other in its community of plants and animals, slope, exposure, soil quality, and climate (Berry 2015)—the forests of the Northeast have become simplified and homogenized.
We have a tendency to take the present look of things to be normal (Berry 2015). Maloof (2016) refers to this as “ecological amnesia.” This shifting sense of “normal” results in a slow diminishment of forest biodiversity. Today, wilderness areas provide the only opportunities to see not only what forests were, but what they could be. Without these ecological baselines we cannot learn the full living language of forests—whether through discovery of the countless species that live there, understanding how a healthy forest functions and responds to a changing climate, or learning how to manage forests in a truly sustainable way. Part of this amnesia stems from the Euro-American tendency to separate humans from the natural world and to compartmentalize and fragment what we study, use, and exploit. Society at large has lost the foundational understanding of interconnectedness in time and space that is a basic truth for most indigenous peoples (Mitchell 2018).
Figure 6 – Arthropods may receive less conservation attention than charismatic animals like birds, fish, and mammals, but they are just as integral to the web of life. This six-spotted tiger beetle was seen at Northeast Wilderness Trust’s Lone Mountain Wilderness Sanctuary in Maine. Photo by Shelby Perry

Figure 7 – Rattlesnake plantain is a type of orchid. Once common across the Northeast, they are now an unusual find. This one was photographed at Northeast Wilderness Trust’s Jack & Margaret Hoffman Wilderness Sanctuary in New Hampshire. Photo by Shelby Perry
A Way Forward
We now face a biodiversity crisis—with species going extinct at 1,000 times the “background” rate (De Vos et al. 2014). The “background” extinction rate is the average number of extinctions that naturally occur without human influence or mass-extinction events. This is often approximated at 1 extinction per million species per year, though recent research suggests the background extinction rate is likely much lower, at around 1 extinction per 10 million species per year.The International Convention on Biological Diversity is finalizing a strategy known as 30 by 30 to halt the decline and extinction of species and allow ecosystems to recover by 2050. The goal is to conserve 30% of every type of ecosystem by 2030 and continue to improve the extent, quality, representation, and resilience of natural and near-natural ecosystems until extinction rates fall to background levels (Convention on Biological Diversity 2021). Included within this goal is a net gain of at least 5% of lands protected as wilderness or ecological reserves by the end of this decade, and a 15% increase by 2050. In the United States, the Biden administration, The Nature Conservancy, Northeast Wilderness Trust, other conservation groups, and many states are embracing these goals, and are also acknowledging that the human connection to nature—especially between indigenous people and their ancestral lands and waters—has to be restored and honored.
“Understanding the complexity, richness, and resilience inherent in wildlands requires the inclusion of diverse perspectives—from the indigenous view that humans are embedded in the natural world, not separate from it, to what science can teach us, to thinking about how a black-capped chickadee sees a forest.”
Today, intact wildlands, including wilderness and ecological reserves, make up just 12% of the United States and less than 6% of the Northeast (Schlawin 2021).The U.S. Geological Survey’s Protected Areas Database classifies wilderness areas and ecological reserves as Gap Status 1 or 2 lands. Gap 1 lands have protections to keep them permanently in their natural state. Gap 2 lands are kept primarily natural, but some natural disturbances, such as wildfires or floods, may be suppressed (U.S. Geologial Survey 2015). We are losing an estimated 37,000 acres of forestland in New England each year (Olofsson et al. 2016). Currently, many ecosystem types, and their unique complements of species, are not represented in reserves or wilderness areas at all, especially in more developed parts of the Northeast. Those that are represented are often too small to maintain the ecosystem functions they were intended to protect, or are not connected enough to provide resilience in the face of climate change. To meet the 30 by 30 goal in the Northeast we need to protect larger and better-connected tracts of wildlands that are embedded in a matrix of sustainably-managed forests, with a focus on intact and representative examples of all the region’s ecosystems (Anderson et al. 2016).
The Nature Conservancy has identified the central and northern Appalachians as a conservation focus area of highest resilience in North America (Anderson et al. 2016). This focus area ranks highly because of its connected land cover, high biodiversity, the amount of carbon it sequesters, and its complex topography, which offers countless microclimates that provide opportunities for species to move. To protect the biodiversity of the Northeast, envision the Appalachians as the trunk of a tree that connects to branches of conserved landscapes throughout the region, and serves as a major movement corridor as species shift ranges in response to a warming climate. Understanding the complexity, richness, and resilience inherent in wildlands requires the inclusion of diverse perspectives—from the indigenous view that humans are embedded in the natural world, not separate from it, to what science can teach us, to thinking about how a black-capped chickadee sees a forest. A connected network of wildlands is where we can come to the truth of our interconnectedness—a truth that is essential to sustain all lives, for generations to come (Mitchell 2018).
About the Author
JANET MCMAHON is a consulting ecologist for conservation organizations, state agencies, and private landowners. She developed a blueprint for an ecological reserves system for the state of Maine and her thesis, The Biophysical Regions of Maine, is used as a framework for landscape-scale conservation efforts by The Nature Conservancy and others.
NORTHEAST WILDERNESS TRUST conserves forever-wild landscapes for nature and people. Founded in 2002, the Wilderness Trust works across New England and eastern New York. The Wilderness Trust owns Wilderness Preserves and Sanctuaries, and protects land through legal means such as conservation easements. The organization currently safeguards more than 64,000 acres of wildlands in six states. Learn more at www.newildernesstrust.org.
References
Anderson, M. G., A. Barnett, M. Clark, J. Prince, A. Olivero Sheldon, and B. Vickery. 2016. Resilient and connected landscapes for terrestrial conservation. The Nature Conservancy, Eastern Conservation Science, Eastern Regional Office. Boston, MA.
Berry, W. 2015. Our only world. Counterpoint Press, Berkeley, California.
Convention on Biological Diversity. 2021. A new global era framework for managing nature through 2030. First Draft Agreement. UN Environment Program, Convention on Biological Diversity.
Davis, M .B. 1996. Extent and Location. Pages 18–32 in M.B. Davis, editor. Eastern old-growth forests: Prospects for rediscovery and recovery. Island Press, Washington, D.C.
De Vos, J. M., L. N. Joppa, J. L. Gittleman, P. R. Stephens and S. L. Pimm. 2014. Estimating the normal background rate of species extinction. Conservation Biology 29(2):452–462.
Gawler, S. C., J. J. Albright, P. D. Vickery, and F.C. Smith. 1996. Biological diversity in Maine: An assessment of status and trends in the terrestrial and freshwater landscape. Maine Natural Areas Program, Augusta, Maine.
Hagan, J. M. and A. A. Whitman. 2004. Late-successional forest: A disappearing age class and implications for biodiversity. FMSN-soor-2. Manomet Center for Conservation Sciences. Brunswick, Maine.
Haney, J. C. and C. P. Schaadt. 1996. Functional roles of eastern old growth in promoting forest bird diversity. Pages 76–88 in M.B. Davis, editor. Eastern old-growth forests: Prospects for rediscovery and recovery. Island Press, Washington, D.C.
Hanski, I. 2000. Extinction debt and species credit in boreal forests: Modeling the consequences of different approaches to biodiversity conservation. Annales Zoologici Fennici 37:271–280.
Haskell, D. G. 2012. The forest unseen. Viking Penguin, London, England.
Haskell, D. G. 2017. The songs of trees. Penguin Random House, LLC., New York, New York.
Hunter, M. L. Jr. 1990. Wildlife, forests and forestry: Principles of managing forests for biological diversity. Prentice-Hall, Englewood Cliffs, New Jersey.
Lapin, M. 2005. Old-growth forests: A literature review of the characteristics of Eastern North American Forests. Vermont Natural Resources Council, Montpelier, Vermont.
Lorimer, C.G. 1977. The presettlement forest and natural disturbance cycle of northeastern Maine. Ecology 58: 1169-1184.
Maloof, J. 2016. Nature’s temples: The complex world of old-growth forests. Timber Press, Inc., Portland, Oregon.
MDIFW (Maine Department of Inland Fisheries and Wildlife). 2015. Maine’s Wildlife action plan. Maine Department of Inland Fisheries and Wildlife, Augusta, Maine.
Meier, A. J., S. P. Bratton and D. C. Duffy. 1995. Possible ecological mechanisms for loss of vernal-herb diversity in logged eastern deciduous forests. Ecological Applications 5:935–46.
Miller, K. M., B. J. McGill, B. R. Mitchell, J. Comiskey, F. W. Dieffenbach, E. R. Mathews, S. J. Perles, J. P. Schmit, A.S. Weed. 2018. Eastern national parks protect greater tree species diversity than unprotected matrix forests. Forest Ecology and Management 414: 74–84.
Mittermeier, R. A., C. G. Mittermeier, T. M. Brooks, J. D. Pilgrim, W. R. Konstant, G. A. B. da Fonseca, and C. Kormos. 2003. Wilderness and biodiversity conservation. PNAS, September 2, 2003, Volume 100 (18) 10309–10313.
Mitchell, S. 2018. Sacred instructions: Indigenous wisdom for living spirit-based change. North Atlantic Books, Berkeley, California.
Olofsson, P., C. E. Holden, E. L. Bullock, and C. E. Woodcock. 2016. Time series analysis of satellite data reveals continuous deforestation of New England since the 1980s. Environmental Research Letters. IOP Publishing.
Selva, S. B. 1996. Using lichens to assess ecological continuity in northeastern forests. Pp. 35–48 in Eastern old-growth forests: Prospects for rediscovery and recovery (M.B. Davis, Ed.). Island Press, Washington, D.C.
Schlawin, J. 2021. Presentation to the Maine Ecological Reserves Scientific Advisory Council on draft ecological reserves section of a report assessing accomplishments and gaps in Maine land conservation. Maine Natural Areas Program, Augusta, Maine.
Sibley, D. A. 2020. What it’s like to be a bird. Alfred A. Knopf, New York.
Simard, S. W. 2021. Finding the mother tree: Discovering the wisdom of the forest. Alfred A. Knopf, New York.
Simard, S. W., I. U. Beiler, M. A. Bingham, J. R. Deslippe, L. J. Philip, and F. P. Teste. 2012. Mycorrhizal networks: Mechanisms, ecology and modeling. Fungal Biology Reviews 26: 39–60.
Swan, C. M. and J. S. Kominoski. 2012. Biodiversity and ecosystem function of decomposition. In: eLS. John Wiley & Sons, Ltd: Chichester. DOI: 10.1002/9780470015902.a0023601 (March 2012).
Tikkanen, O., P. Martikainen, E. Hyvarinen, K. Junninen and J. Kouli. 2006. Red-listed boreal forest species of Finland: associations with forest structure, tree species, and decaying wood. Annales Zoologici Fennici 48(4): 373–383.
U.S. Department of Agriculture Northern Research Station. 2019. Forest Inventory and Analysis, Maine Forest Inventory tool: http://www.fia.fs.fed.us/tools-data/default.asp
U.S. Geologial Survey. 2015. Enviro Atlas Fact Sheet www.epa.gov/enviroatlas
Whitman, A. A., and J. M. Hagan. 2009. A revised rapid-assessment late-successional index for common northeastern forest types. Forest Mosaic Science Notes 2009-1. Manomet Center for Conservation Sciences, Brunswick, Maine.
Whitman, A. A. and J. M. Hagan. 2000. Herbaceous plant communities in upland and riparian forest remnants in western Maine. Mosaic Science Notes #2000-3. Manomet Center for Conservation Sciences, Brunswick, Maine.
Zlonis, E.J. and G.J. Niemi. 2014. Avian communities of managed and wilderness hemiboreal forests. Forest Ecology and Management 328: 26–34.
Additional References
Department of Environmental Conservation. 2015. New York State Wildlife Action Plan. New York
Department of Environmental Conservation, Albany, N.Y.
Johnson, E.A. and D. Smith, eds. 2006. Legacy: Conserving New York State’s Biodiversity. American Museum of Natural History, New York State Biodiversity Research Institute, New York State Department of Environmental Conservation, New York Natural Heritage Program and The Nature Conservancy, Albany, N.Y.
Massachusetts Division of Fisheries and Wildlife. 2015. Massachusetts State Wildlife Action Plan 2015. Westborough, Massachusetts.
Terwilliger Consulting Inc. 2015. 2015 Connecticut Wildlife Action Plan. Prepared for The Nature Conservancy and the Connecticut Department of Energy and Environmental Protection, Bureau of Natural Resources.
Terwilliger Consulting Inc. 2015. 2015 Rhode Island Wildlife Action Plan. Prepared for The Nature Conservancy and the Rhode Island Department of Environment Management Division of Fish and Wildlife.
Vermont Fish & Wildlife Department. 2015. Vermont Wildlife Action Plan 2015. Vermont Fish & Wildlife Department. Montpelier, VT. https://vtfishandwildlife.com/node/551
Read Next

What is visitor use management? Identifying functional and normative postulates of an interdisciplinary field of study
The axioms of Visitor Use Management can guide us towards a future that enriches the lives of the public while preserving our cherished resources for generations to come.

Are We in Need of “Triage” for Wilderness?
As part of a larger management strategy, “triage” actions can work with long term decision-making to provide wilderness managers with the best practices necessary for wilderness stewardship.

Nature in the Anthropocene: What it no longer is, Will never again be, and What it can become
Nature changes in response to changing conditions, and so must our conception of it.